研究テーマ
-
ダイズの裂莢性
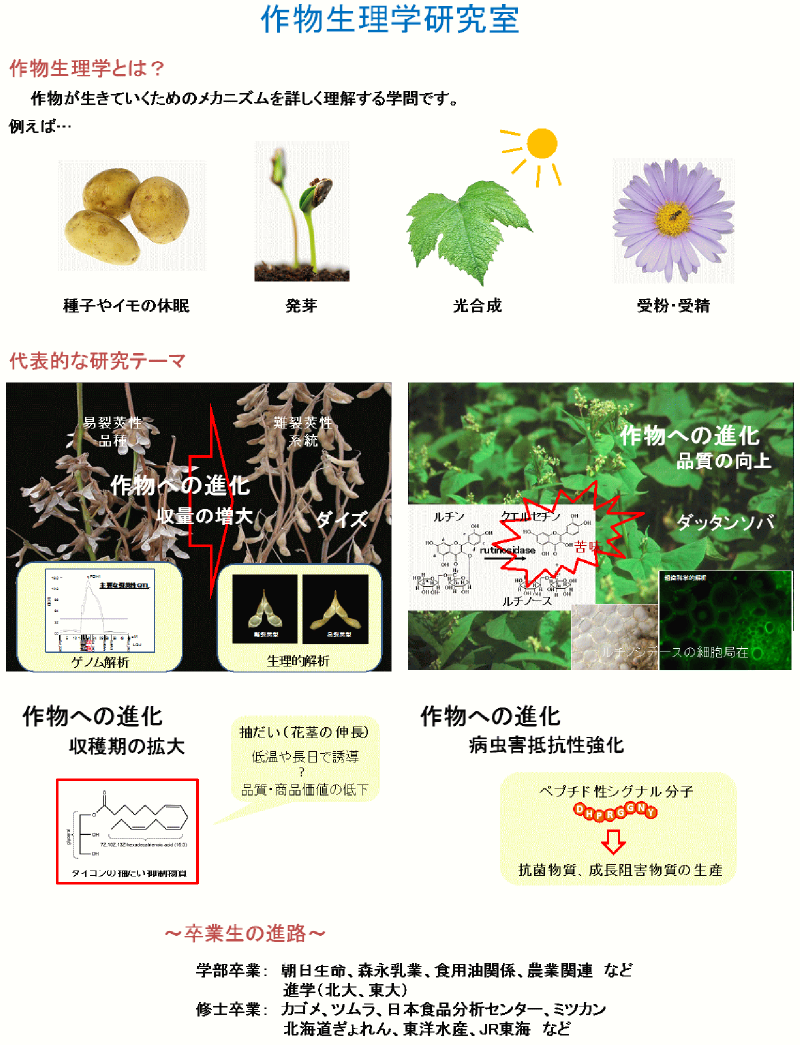
莢の裂けやすさ(裂莢性)は種子の落下のしやすさ・分布を決定づける性質の一つです。ダイズにおいては、早期の種子落下による減収を防ぐため、難裂莢性系統が選抜・育種されてきました。裂莢は種子・莢の乾燥に伴い生じますが、低湿度下においても難裂莢性系統の裂莢は易裂莢性系統のそれよりも著しく遅れます(図1)。
他機関と当研究室の共同研究により、裂莢性に高い効果を持つ遺伝子としてPDH1が見出されています(図2)。PDH1はdirigent-likeタンパク質ファミリーの一つをコードし、細胞壁の性質を変化させると考えられていますが、その機能については不明な点も多くあります。当研究室ではPDH1について詳細な解析を進めています。

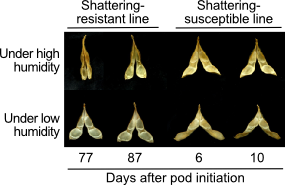
図1. ダイズの難裂莢性系統と易裂莢性系統
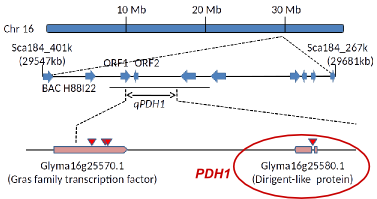
図2. PDH1のマッピング
参考論文:
Funatsuki H ... and Fujino K. Molecular basis of a shattering resistance boosting global dissemination of soybean. Proc Natl Acad Sci U S A. 2014. 111: 17797-17802. DOI: 10.1073/pnas.1417282111. PMID: 25468966.
Funatsuki H ... and Fujino K. Mapping and use of QTLs controlling pod dehiscence in soybean. Breed Sci. 2012. 61: 554-558. DOI: 10.1270/jsbbs.61.554. PMID: 23136494.
-
ダッタンソバの苦味
ダッタンソバはタデ科ソバ属の一年草であり、高い健康効果を持つルチンという物質を多く(フツウソバの50-100倍)含むことから、ソバの原料として利用されています(図3)。しかし、製麺過程で細胞が破砕されるとルチノシデースという酵素が細胞外に流出し、ルチンをケルセチンという苦味の強い物質に変換するため、従来のダッタンソバ品種を用いたソバ麺は食味が悪いとされてきました。
近年、苦味の軽減された「満天きらり」というダッタンソバ品種が開発されました。当研究室では、従来品種と「満天きらり」のルチノシデース遺伝子について詳細な比較解析を行っています(図4)。また、その成果を利用して、製麺過程での満天きらりの純度(他品種の混合率)を検定するためのDNAマーカーを開発しています。

図3. フツウソバ(左上)とダッタンソバ(左下、右)
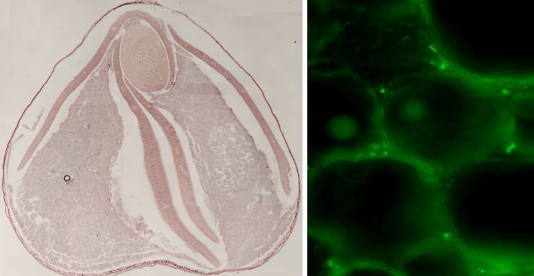
図4. ダッタンソバ種子におけるルチノシデース遺伝子の発現部位(左図赤色部分)とルチノシデースの免疫染色(右図輝点)
参考論文:
Suzuki T et al. Breeding of 'Manten-Kirari', a non-bitter and trace-rutinosidase variety of Tartary buckwheat (Fagopyrum tataricum Gaertn.). Breed Sci. 2014. 64: 344-350. DOI: 10.1270/jsbbs.64.344. PMID: 25914589.
-
アスパラガスの雌雄性
食用アスパラガスは、個体ごとに雌花を着けるもの(雌株)と雄花を着けるもの(雄株)に分かれるという性質(雌雄異株性)を持ちます。雌株は雄株よりも可食部の若茎の成育が不揃いであり、また雌株に着いた種子が落下・発芽するとその個体が雑草化して栽培管理上不都合が生じるため、農業生産には雄株が好んで用いられます。雄株には交配後代に雄株のみを生じさせるものがあり、そのようなものを超雄性株と呼びます。超雄性株は栽培用種子の生産や育種に必須です。
アスパラガスの雌花においては、雄蕊が分化後成育を停止し退化します。また雄花においては、雌蕊が分化後成育を停止します。これが雌・雄花の形態の差異を生みます(図5)。当研究室では、この過程における細胞死の動態や、植物ホルモンの影響について解析を進めています。
アスパラガスの雌雄性はMと呼ばれる遺伝子座により支配されることが知られています。すなわち、遺伝子型mmの株が雌株、Mmの株が雄株、MMの株が超雄性株です。当研究室では、M座遺伝子の候補を単離し(図6)、その機能の解析も進めています。

図5. アスファルトに生えるアスパラガス(左)とアスパラガスの雌・雄花(右)
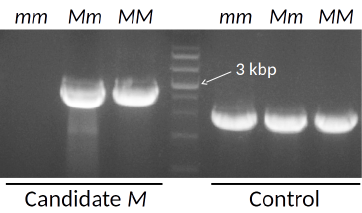
図6. 雌株(mm)、雄株(Mm)、超雄性株(MM)のゲノムを鋳型として用いたPCRによるM座遺伝子の候補の検出
参考論文:
Harkess A et al. Sex-biased gene expression in dioecious garden asparagus (Asparagus officinalis). New Phytol. 2015. 207: 883-892. DOI: 10.1111/nph.13389. PMID: 25817071.
Horiuch K, Adachi Y, Kasai N, Yamagishi M, Masuda K. Identification of homozygous male plants by quantitative analysis of a nucleotide sequence linked to the sex-determination locus in Asparagus officinalis L. Journal of the Japanese Society for Horticultural Science. 2011. 80: 308-313. Full text.
Asada Y, Kasai N, Adachi Y, Kanno A, Ito N, Yun PY, Masuda K. A vegetative line of asparagus (Asparagus officinalis) with a homeotic change in flower development is correlated with a functional deficiency in class-B MADS-box genes. Journal of Horticultural Science & Biotechnology. 2006. 81: 874-882. DOI: 10.1080/14620316.2006.11512153.
Jamsari A et al. BAC-derived diagnostic markers for sex determination in asparagus. Theor Appl Genet. 2004. 108: 1140-1146. DOI: 10.1007/s00122-003-1529-0. PMID: 15067401.
-
植物の核内膜構成タンパク質
動物細胞の核膜は、脂質二重膜、膜内在性タンパク質、核膜孔複合体、ラミナにより構成されています。ラミナは脂質二重膜の核質側において網目状に配向する繊維状の構造体(中間径フィラメント)であり、ラミンというタンパク質が重合することにより形成されます。植物は、ラミンと似たアミノ酸配列を持つタンパク質は持ちませんが、核にラミナ様の構造体を持ち、これの構成因子としてNMCPというタンパク質が見出されています。NMCPはラミンと起源を異にすると考えられますが、ラミンと似た高次構造を取ると予測されています(図7)。ラミナやラミナ様構造は核の形態や機能を維持するのに必要であり、ラミンやNMCPにおけるある種の変異は、深刻な疾患や成育障害を引き起こします。
当研究室では、NMCPの可視化(図8)やNMCP結合タンパク質の探索を行っています。植物細胞の核の構成因子の動態・機能には未だに不明な点が多くありますが、NMCPとその結合因子を詳細に解析することでそれらを明らかにしたいと考えています。
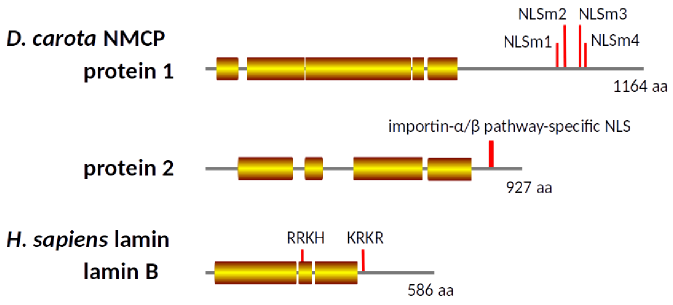
図7. NMCP1(Protein 1)、NMCP2(Protein 2)、ラミンBの二次構造予測(coiled-coil構造を取りうる領域を色付きの四角形で示す)
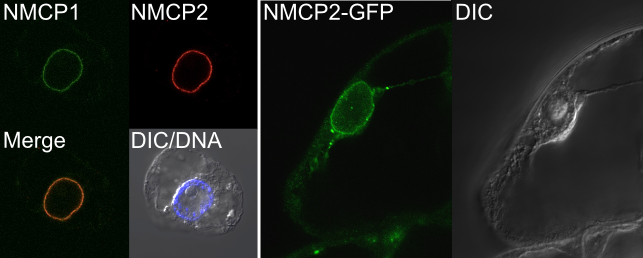
図8. 蛍光免疫染色法によるNMCP1とNMCP2の検出(左)とNMCP2-GFP融合タンパク質の細胞内局在(右)
参考論文:
Kimura Y, Fujino K, Ogawa K, Masuda K. Localization of Daucus carota NMCP1 to the nuclear periphery: the role of the N-terminal region and an NLS-linked sequence motif, RYNLRR, in the tail domain. Front Plant Sci. 2014. 5: 62. DOI: 10.3389/fpls.2014.00062. PMID: 24616728.
Ciska M, Masuda K, Moreno Díaz de la Espina S. Lamin-like analogues in plants: the characterization of NMCP1 in Allium cepa. J Exp Bot. 2013. 64(6): 1553-1564. DOI: 10.1093/jxb/ert020. PMID: 23378381.
Kimura Y, Kuroda C, Masuda K. Differential nuclear envelope assembly at the end of mitosis in suspension-cultured Apium graveolens cells. Chromosoma. 2010. 119: 195-204. DOI: 10.1007/s00412-009-0248-y. PMID: 19997923.
Masuda K et al. Peripheral framework of carrot cell nucleus contains a novel protein predicted to exhibit a long alpha-helical domain. Exp Cell Res. 1997. 232: 173-181. DOI: 10.1006/excr.1997.3531. PMID: 9141634.
Nomura K, Saito W, Ono K, Moriyama H, Takahashi S, Inoue M, Masuda K. Isolation and characterization of matrix associated region DNA fragments in rice (Oryza sativa L.). Plant Cell Physiol. 1997. 38(9): 1060-1068. PMID: 9360323. Full text.
Masuda K, Takahashi S, Nomura K, Arimoto M, Inoue M. Residual structure and constituent proteins of peripheral framework of the cell nucleus in somatic embryos from Daucus carota L. Planta. 1993. 191: 532-540. DOI: 10.1007/BF00195755.
-
機械的刺激の感知の分子機構
植物は接触などの機械的な刺激を感知し、それに対して適応的な応答を行いますが、その分子機構には不明な点が多くあります。これについて新たな知見を得るべく、当研究室ではVIP1というタンパク質に着目しています。VIP1はシロイヌナズナの転写因子(他の遺伝子産物の量を調節するタンパク質)であり、機械的刺激を細胞が感受すると、VIP1の細胞内存在部位は一過的に細胞質から核へと変化します(図9)。遺伝子工学的な手法により、VIP1-SRDXoxという変異型のVIP1を用いてVIP1の機能を撹乱すると、根の接触屈性(根端が物体に触れた際にそれを避けるように屈曲する性質)が増大し、根が大きく波打って成長します(図10)。これにはオーキシンという植物ホルモンが関わるということが示唆されています。
当研究室では、VIP1の細胞内存在部位の制御機構やVIP1の生理的な役割などについて更に解析を進めています。VIP1を足掛かりに、機械的刺激の感知に関わるタンパク質を更に多く同定していきたいと考えています。
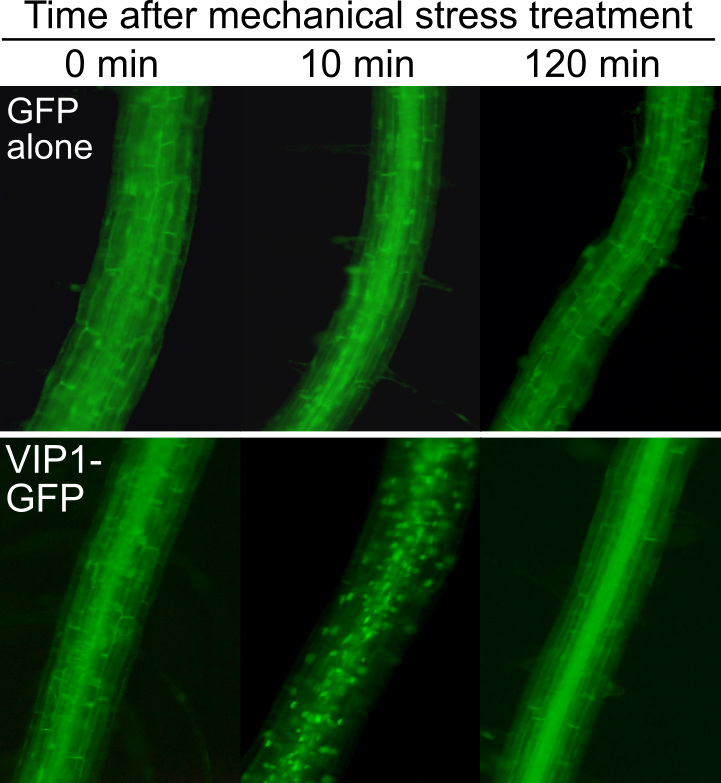
図9. 機械的刺激を受けた根の細胞におけるVIP1の核移行(下段中央のドットが核に相当)

図10. 接触刺激を与えながら根を成育させた際の根の波打ち(左)の程度は、VIP1-SRDX過剰発現体(VIP1-SRDXox)において大きい(右)
参考論文:
Tsugama D, Liu S, Takano T. VIP1 is very important/interesting protein 1 regulating touch responses of Arabidopsis. Plant Signal Behav. 2016. 11(6): e1187358. DOI: 10.1080/15592324.2016.1187358. PMID: 27171129.
Tsugama D, Liu S, Takano T. The bZIP protein VIP1 is involved in touch responses in Arabidopsis roots. Plant Physiol. 2016. 171(2): 1355-1365. DOI: 10.1104/pp.16.00256. PMID: 27208231.
Tsugama D, Liu S, Takano T. Analysis of functions of VIP1 and its close homologs in osmosensory responses of Arabidopsis thaliana. PLoS One. 2014. 9(8): e103930. DOI: 10.1371/journal.pone.0103930. PMID: 25093810.
Tsugama D, Liu S, Takano T. A bZIP protein, VIP1, is a regulator of osmosensory signaling in Arabidopsis. Plant Physiol. 2012. 159(1): 144-155. DOI: 10.1104/pp.112.197020. PMID: 22452852.
講座(研究室)の歴史
-

1908 研究室の原型である植物学第二講座(植物生理学講座)が設立される。
東京帝国大学より招聘された柴田桂太が初代教授に就く。~1918 大野直枝、群場寛が教授を務め、植物生理学の基礎を築く。 1919 坂村徹が教授となり、植物生理学の集大成として「植物生理学」上下2巻を著す。
坂村に師事していた木原均は、後に世界的に有名なコムギゲノム研究を展開する。戦後 混乱と食糧不足が続く中、リュックを担いでバレイショを集め、バレイショの生理についての研究を進める。
新設された理学部植物生理学講座と区別するため、講座名が「作物生理学講座」に変更される。1955 バレイショの成育・休眠の機構に関する研究の成果により、教授・田川隆が日本作物学会賞を受賞する。 1970 ジベレリンの減少が塊茎形成の必要条件であることを明らかにした教授・岡沢養三が日本作物学会賞を受賞する。 ~1997 教授・喜久田嘉郎がイネ、ソバ、牧草なども研究材料として取り入れる。また、研究に分子生物学的研究手法を積極的に取り入れ、バレイショの塊茎組織や葉肉プロトプラストから個体を再生する系を確立する。
大学院重点化に伴う組織改革により、作物学講座、植物病理学講座、作物栄養学講座と共に作物生産生物学大講座を形成する。2001 教授・幸田泰則が「バレイショの塊茎形成誘導物質・ジャスモン酸類似物質の発見」により日本作物学会賞を受賞する。 ~2013 ダイコン、ホウレンソウなど、更に多くの作物を研究材料とするようになる。
組織改革により、園芸学講座、作物学講座、植物病理学講座と共に作物生産生物学大講座を形成する。~2017 ニンジン体細胞からの胚形成の機構や新規植物細胞骨格タンパク質NMCPを発見した増田清が教授を務めた。