次のようなテーマに興味を持ち、研究を行ってきました。
・ ゴール形成アブラムシの多様性とゴール寄生性の進化
・ゴール形成アブラムシの生物地理;隔離分布と進化の停滞
・ゴール形成アブラムシのゴール形成場所選択
・ゴール形成アブラムシの局所的適応・ホストレース形成
・寄主植物の多様性とゴール形成アブラムシに働く選択圧
・近親交配がアブラムシの発生に与える影響;隠蔽されたトレードオフ
・ゴール形成アブラムシにおける跳躍的種分化
・兵隊アブラムシの進化条件:コンピューターシミュレーション
・サッポロフキバッタの地理的変異と種形成
・アブラムシの分類
○ ゴール形成アブラムシの多様性とゴール寄生性の進化
ニレハマキワタムシ属(Eriosoma属(写真あり))のゴール形成アブラムシでは、寄主植物(ハルニレ)上で6種ほどの同属種が共存しています。どの種もよく似た葉巻型のゴールをハルニレの葉に形成します。ある種のEriosomaが作ったゴールの中に同種あるいは他種のEriosomaが侵入し、ゴール内で共存することがしばしば起こります。こうした入り込みは偶発的に生じることで、適応的な行動というわけではありません。しかし、ある種のEriosoma(ヤドリハマキEriosoma yangi parasiticum)は、自分ではゴールを作らずに、もっぱら他種のゴールに侵入する性質を進化させました。この侵入者は、他種のゴールに侵入後、ゴール形成者を殺し、ゴールを乗っ取って自分の子供を産みます。こうした寄生はスズメバチやマルハナバチに見られる社会寄生性や、あるいはカッコウの托卵とよく似た現象です。どのようにして、こうした性質が進化したのでしょう。
寄生性を示すヤドリハマキは、本来ハルニレにいた種ではなく、外来の種であることがわかりました。ハルニレの近縁種にアキニレという植物があります。ハルニレが比較的冷温帯に分布するのに対し、アキニレは東京以西に分布しています。ヤドリハマキの祖先型はアキニレを本来の寄主にしていました。ヤドリハマキの亜種
アキニレハマキワタムシ Eriosoma yangi yangiは、アキニレで葉巻型のゴールを形成しています。他種ゴールへの寄生性は一部の集団がアキニレからハルニレへの寄主の乗り換えに伴って進化したと考えています。
ゴールを形成する昆虫は寄主特異性がたいへん高いのが特徴です。その結果、特定の植物にしかゴールを作ることができません。ところが、アブラムシの有翅型は、間違えて近縁他種の植物を寄主として選び、そこに子孫を産み付けることがしばしばあります。孵化した子孫は、寄主ではない植物にゴールを作ることはできません。しかし、そこに近縁種のゴールがあれば、ゴールの中で生育することは可能です。いったん、誤った寄主選択が行われると、その寄主上で生き残るためには、他種のゴールへ侵入するしかありません。こうしてゴール侵入には強い選択が働き、やがてはゴール形成者を殺してしまう性質も獲得したのだと考えています。
したがって、Eriosoma yangiという種は2つのタイプに分かれます。1つはアキニレでゴール形成をする祖先的タイプと、もう一つはハルニレで他種のゴールに寄生して生きているタイプです。生活史は大きく異なりますが、形態的には若干の違いが見られるだけです(寄生タイプの方が活動に適した形態を示します)。この2つの集団をどのように扱うのが適切なのでしょうか?交配実験が容易には行えない生物では、客観的に種の区分けを行うことは困難です。現在は、伝統的な形態的分類学の立場に立って、明確な形態的不連続性が認められないこれらの分類群を同一種として扱い、ヤドリハマキとアキニレハマキは亜種と見なしています。
「ヤドリハマキは本来ハルニレにいた種ではなく、外来の種」と上に書きましたが、このことは観察結果から分かったわけではありません。寄生性のアブラムシが進化した原因をあれこれ考えていたときに、どうしても寄生性を進化させる有利性が思いつかず、困り果てていたときに、ある日だしぬけに「思いついた」ことなのです。この寄生者の祖先はよそからやってきたに違いないと思いつき、そうした観点でアキニレの分布地域まで調べに行ったら、まさしくそこに祖先がいたのでした。
○ ゴール形成アブラムシの生物地理;隔離分布と進化の停滞
ゴール形成アブラムシは特定の寄主植物に特殊化しており、寄主植物とともに分布を広げてきました。Eriosomatini(ワタムシ族)のアブラムシは、基本的にはニレ科植物にゴールを形成します。ニレ属にゴールを形成するスゲワタムシColopha属はユーラシアと北米東部に分布しています。この属は6種からなる小さなグループで、うち3種は単為生殖だけで増殖します。有性生殖を行う3種はゴール形成者で、ニレ属のUlmus
laevis とUlmus americanaに寄生します。ニレ属のなかでU. laevisとU. americanaは最も近縁で、ヨーロッパと北米東部に隔離分布しています。有性生殖種のColophaも寄主とともに隔離分布します。寄主植物は、アジアには現在分布していませんが、第三紀中新世中期ごろまではヨーロッパーアジアー北米に広域分布しており、アジアではその要素が絶滅したと考えられます。実際、Ulmus
americanaに近いと思われるニレの化石が日本の中新世の地層から得られています。
U. americanaグループが絶滅した東アジアでは、単為生殖種のColopha kansugeiが2次寄主のスゲ上に分布します(本種写真)。この種は、かつては1次寄主のニレにゴールを作っていたと考えられますが、1次寄主の絶滅後、2次寄主上に取り残され、以後単為生殖を長期にわたって続けてきたと考えられます。Colophaの他の2種の単為生殖種もこうした、「遺存種」だと考えています。単為生殖3種の分布は、第四紀のリフュージであった地域に含まれます。C.
kansugeiは本州西部、南西諸島、台湾、中国南部、タイ北部、ネパールに至る広大な地域に分布し、集団間で遺伝的に分化していますが、同一種と見なされています。これほど広い地域に分布しながら、同一種でいられるのは単為生殖に原因があるのかもしれません。形態的に不連続性で区切られた存在、いわゆる「種」が生じるのは、有性生殖が存在するからではないかと考えています。
○ ゴール形成アブラムシのゴール形成場所選択
閉鎖型ゴールを形成するアブラムシは、ゴールを解剖することによって簡単に子孫の数を知ることができるために、ゴール形成場所選択と適応度の関係を明らかにするための研究材料としてしばしば使われてきました。これまでの有名な研究(Whitham, 1978)では、ゴール形成アブラムシは大きな葉を選んでゴールを形成すると多くの子供を残せることが報告されています。1枚の葉の中でも、基部に近い側にゴールを作ると、適応度はさらに高まります。ところが、この結果は私の調べたゴール形成アブラムシTetraneura sp.(いまだ未記載)には当てはまりませんでした。Tetraneuraでは、ゴール内で生まれる幼虫の数は葉のサイズやシュートのサイズとは何の関係もありません。Tetraneuraでは、Whithamの結果とは逆に、葉の先端部にゴールを形成した方が適応度が高いことも明らかになりました。Tetraneura幹母がより多くの子供を残すには、場所の選択よりも、早く孵化して早い時期に成長している葉を刺激することが重要でした。つまり、種によって、自然選択のかかり方がまったく異なっているのです。当初は、春早く成長を始める植物組織はきわめて栄養条件が良く、遅れるにしたがって栄養条件が悪くなると考えていました。ところが最近、これまでの解釈は、おそらくWhithamのものも含めて、まったく見当違いであったことが判明しました。ここから先はまだ発表していませんが、簡単に言えばゴール内で生まれる子供の数はゴール場所選択の結果決まるというわけではなく、かなりの部分が初めから決まっていたのです。ゴールアブラムシの卵サイズには種内でもかなりの変異があります。大きな卵ほど早く孵化が起こります。さらに、大きい卵からは大きな幼虫が孵化してきますが、大きな幼虫は孵化した時点でよりサイズの大きな生殖腺を持っています。その結果、早くに孵化した大きな幼虫は良い場所を占め、多くの子供を残すことになります。産子数は、アブラムシの場合母性効果を強く受けます。したがって、ゴールを採集して幼虫数を数えただけでは、自然選択の働きは明らかにはならないのです。Whithamの示した例は、寄主植物の効果によって植食性昆虫の適応度が大きく左右されるという見方をもたらしましたが、実際にはゴール形成者の個体差(遺伝的な側面も含む)が適応度に大きな影響を与えていました。
○ ゴール形成アブラムシの局所的適応・ホストレース形成
ハルニレにゴールを作るニレイガフシアブラムシ(Kaltenbachiella japonica)は寄主転換せず、周年ハルニレ上で生活を送ります。このアブラムシは寄主の各個体の性質に適応し、同じ寄主集団内で遺伝的に分化を遂げていることが明らかになりました。つまり、ハルニレの一本一本に対して局所的適応を遂げた固有の遺伝的集団が作られているのです。これは、微少なレベルでも遺伝的分化が進むことを示すよい例となるでしょう。こうした遺伝的分化が進むのは、寄主のハルニレが多様な表現型(個性)を示すことと、ニレイガフシアブラムシが移動力を欠いているためです。ハルニレは、春に芽吹き時期に木によって大きな違いが見られます。北大構内でも、早く芽吹く木と遅く芽吹く木では最大で3週間くらいのずれがあります。ニレイガフシアブラムシは、ハルニレの芽吹きと同時に越冬卵から孵化しゴール形成を始めないとうまくゴールが作れません。こうしたシンクロナイゼーションの必要性から、卵の最適な孵化時期はハルニレ個体ごとに異なります。興味深いことに、同じ温度条件に置いて越冬卵を孵化させたところ、早く芽吹き木から取った卵は、遅く芽吹く木から取った卵よりも平均的に早く孵化することがわかりました。すでに遺伝的な分化が生じていたのです。量的遺伝学の手法を用いると孵化時期の分散を、1)ハルニレ個体間、2)ハルニレ個体内のゴール間、3)ゴール内に分割することができます。最も大きな分散成分が見られたのはハルニレ個体間でした。この結果も、局所的な遺伝的分化を支持しています。
○ 寄主植物の多様性とゴール形成アブラムシに働く選択圧
上記の局所的遺伝的分化は移動能力の低いアブラムシに見られた現象です。では、ハルニレにゴールを形成するアブラムシのなかで、高い移動能力を持つ種類ではどうなるのでしょうか?この問題を、寄主転換を行う(すなわち移動能力が高い)アブラムシで調べてみたところ、予想通り局所的遺伝的分化は生じていませんでした。ゴールを作るTetraneuraの一種は、芽吹きの早い木でも遅い木でもお構いなしに産卵してしまいます(何か寄主選択の基準があるのでしょうがよくわかっていません)。したがって、芽吹きの早い木ではアブラムシの孵化は相対的に遅れることになります。逆に芽吹きの遅い木ではアブラムシは相対的に早く孵化します。このために、シンクロナイゼーションがうまくいかず、多くの孵化幼虫が選択的死亡を受けていました。しかし、その一方で、このTetraneuraには孵化時期に関して多量の遺伝分散が保存されていることがわかりました。半兄弟分析を行うと、卵孵化時期の遺伝率はたいへん高く75%ほどと推定されました。環境が異質で、しかも生物がどの環境に直面するかわかっていないと、生物は「特定の環境」に対して適応することができません。どの環境に対しても、中途半端な適応しか示せないのです。面白いことに、こうした環境で、遺伝分散はたいへん高い状態に保たれていました。もし環境が安定していたら、最適な遺伝子型だけが占めてしまい、遺伝分散は小さい状態に保たれると考えられます。
○ 近親交配がアブラムシの発生に与える影響;隠蔽されたトレードオフを明らかにする
近親交配をさせると形態形質に大きな変化が生じることをユキムシを使って調べています。Drosophilaを使ったこれまでの報告では、近親交配は形質の平均値を数%変化させるということですが、アブラムシでは極めて大きな変化が一部の家族に生じました。ユキムシの場合、卵サイズを調べ、卵から孵化する幼虫のサイズを測定しているので、野外では死亡するような個体の形態も調べられます。卵のサイズが一定でも、近親交配させた場合には、孵化する幼虫のサイズにバラツキが見られました。資源が限られているわけですから、このようなことは考えにくいことですが、体サイズが大きくなる分、生殖腺の発達が悪くなっていました。本来、外胚葉(体表)と内胚葉(生殖腺)のレベルでトレードオフが存在していると思われます。しかし、遺伝子が正常に働く場合にはトレードオフが存在することが隠蔽されています。発生のすすみには極めて強い安定化選択が働くからです。近親交配を強制すると、普段は隠されている劣性遺伝子の効果が現れ、発生経路が乱れた個体が生じてくるのです。近親交配を強制することで、表現型の分散を大きくすることが可能で、その結果普段は目にできないトレードオフを明らかにすることができます。
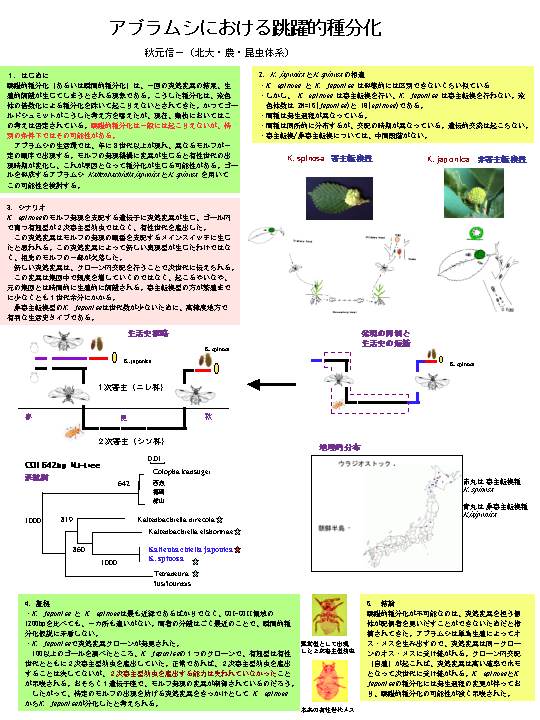
○ ゴール形成アブラムシにおける跳躍的種分化:寄主転換性
アブラムシから非寄主転換性アブラムシの瞬間的種分化
跳躍的種分化というのは、Noe-Darwinismの枠組みの中では最もタブーに近いものです。これまで、跳躍的種分化の例は染色体の倍加に伴う種分化以外には確実な証拠がありません。
タブーに挑戦するつもりはないのですが(わたしもNoe-Darwinismの枠組みの中で仕事をさせてもらっています)、「たまたま」跳躍的種分化の事例が見つかってしまいました。もちろん、これが生じるには特別の条件が必要です。突然変異体が単為生殖によって個体数を増殖させる過程を持たない生物では、跳躍的種分化は生じ得ないでしょう。
ゴール形成アブラムシでは寄主転換を行う種が多く、そうした種では樹木の1次寄主から草本類の2次寄主へと初夏に有翅型が移動し、秋には再び2次寄主から1次寄主へと有翅型が戻ってきます。アブラムシは、単為生殖によって年に何世代も繰り返し、秋に1次寄主上で有性生殖を行います。
見いだされた事例というのは次のようなものです。ハルニレにゴールを作り、寄主転換を行うニレオオイガフシという種類と、それと形態が全く区別のできないニレイガフシアブラムシのペアーが同所的に見つかりました(札幌市の周辺です)。ニレオオイガフシでは、ゴールから現れる有翅型はお腹に単為生殖を行う幼虫を持っており、2次寄主へ移動して産子します。一方、ニレイガフシでは、ゴールから出現する有翅型はお腹に有性生殖を行う幼虫を持っており、寄主転換せずにハルニレに残り、ハルニレの枝で有性世代を生んでしまうのです。この2種は発生過程に大きな変化が生じており、一方では有翅型のお腹で単為生殖幼虫が育ち、他方では有性生殖幼虫が育ちます。両タイプの幼虫は形態が全く異なります。寄主転換するニレオオイガフシは秋に有性生殖が起こり、非寄主転換タイプのニレイガフシは夏に有性生殖を行うので交配のチャンスはありません。また両種は染色体数が異なります。
ここで起こった変化は世代の短絡です。
A→B→C→D とモルフが出現していた祖先タイプから(Dが有性世代)
A→B→D というタイプが生じたために、元のタイプとは交配ができなくなりました。2つのタイプの間に中間状態はありません。どうしても、新しいタイプの方が1世代少なくなります。したがって、生じたときには、一気に生殖隔離が確立したと考えられます。
こうした変化が起こったことは形態と生活史の比較によって前から気づいていましたが、最近DNA配列の比較と突然変異個体の発見によって、この可能性はますます高まりました。DNA配列では両種は全く違いがありません。また、突然変異というのは、ニレイガフシ(A→B→Dの種)の中に、失われたはずのモルフCが異常個体として出現したのです。まさに先祖帰り的な変化です。高木先生、ついにatavismがみつかりましたよ!
○ 兵隊アブラムシの進化条件:コンピューターシミュレーション
兵隊アブラムシが私のお師匠さんの青木重幸さん(現、立正大学)によって発見されました。どのような生態条件の時に兵隊(コロニー防衛)が進化しやすいのかをシミュレーションで調べたのがこの研究です。昔のことなので、メインフレームコンピューターを使ってFortranでがんがん計算しました。
兵隊アブラムシというと通常幼虫と不妊兵隊の2型を示す例が有名ですが、もっと原始的なタイプでは、すべての1令幼虫が捕食者に対して攻撃性を示し、専門の兵隊を持ちません。他の齢の幼虫および成虫は全く防衛に参加しません(いわば、国民皆兵型です)。国民皆兵型の1令幼虫の場合は、捕食者にやられなければ、その後も成長していけます。このような種(実例)では、1齢幼虫は他の齢に比べて幼虫期間が長いことがわかっていました。1令期間が長いほど、そのコロニーでは防衛個体の比率が高くなります。こうした原始的なタイプを想定してシミュレーションを行いました。つまり、どういう条件の下で、長い1齢期間(すなわち高い比率の防衛個体)が最適になるかを調べたわけです。この研究は、人間社会で最適徴兵期間を決めるのと同じですね(これって、軍事研究だったのか?)。
一定期間内にコロニーサイズを最大化するという目標を設定すると、捕食さえなければ、世代時間をできるだけ短くして何世代も繰り返せば、増殖率は高まります。しかし、捕食者に対して防衛する場合には、防衛個体をある程度確保しなければいけないので、1令期間が長くなります。これによって全体の世代時間は延び、増殖率は低くなります。防衛効果と増殖率の間にトレードオフが現れるわけです。
捕食圧が高いと、シミュレーションではおもしろい最適解が現れました。春から夏にかけて、成虫は子供を産んでいくわけですが、生まれた幼虫は成長を止めてしまって1令のままとどまるのです。つまり、成虫以外のコロニーのすべてが1令幼虫(防衛個体)という状態が秋の最終時点直前まで続くのです。そして、最後の最後になって1令幼虫は一気に成長を再開し、最終時点に滑り込むようにしてすべてが成虫になります。つまり、春から秋まで、世代を全く回さずに、コロニーを重武装して捕食者から守りきるという戦略が理論上はあり得るのです。
シミュレーションをしていた時点では、まさかこんなアブラムシがいようとは思いませんでした。ところが、やがて北米でこのようなアブラムシが実際に見つかったときには、心底驚き、こうしたシミュレーションでも新たな認識を得られることがわかり、たいへんうれしくなりました。
○ サッポロフキバッタの交尾行動 (ref.60)
サッポロフキバッタは北海道各地で遺伝的分化をとげた集団から成り立っていますが、ある集団のオスは交尾活力が高く(ガツガツしている)、逆にメスはオスの交尾を強くはねつけます。別の集団では、オスもメスもお上品で、おとなしめです。こうした集団が出会うと、非対称的な交尾頻度が観察されます。つまり、2集団からオスメス同数を選んできて飼育容器にいれると、交尾活力の高い集団のオスは、自分の集団のメスとよりも、おとなしい集団のメスと頻繁に交尾します。逆に、交尾活力が低い(おとなしい)集団のオスと交尾拒絶力が高い集団のメスの間ではほとんど交尾がおこりません。こうした、非対称な交尾は、どの2集団を選んでも観察されます。サッポロフキバッタではメスにおいて、交尾拒否行動の強さに集団間で違いがあり、オスの交尾活力は、メスの拒絶力に対抗して進化したと考えています。どうして、このような交尾拒絶がメスで進化してきたのでしょうか?この問題をsexual conflictの概念で説明しようとしています。
○ アブラムシの分類
ここまで読んでくれた人はたぶん「この人はいろいろやっているのだなー」と思ったことでしょう。しかし、私の本当の専門は、アブラムシの分類です(笑)。正しい分類学者を目指して日夜努力中ですが、いろいろなことに手を出しすぎ、記載しなくてはいけないアブラムシが不良債権のごとく蓄積してしまいました。もう処理できん、すべて*野君に丸投げだ!
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}