ホウレンソウに見られる多様な「性」
ホウレンソウ(Spinacia oleracea) は,雌雄異株として一般に知られていますが,実際には雄株および雌株の他に,様々な品種,系統および交雑後代から雌雄両方の機能を備えた間性株(雌雄同 株)が見出されます. このページでは、形態的特徴からホウレンソウの性について解説します。
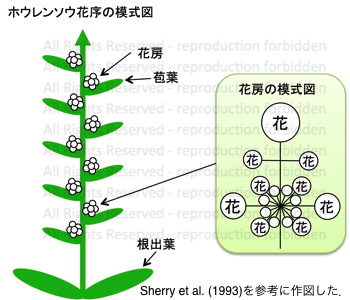
ホウレンソウの花序
ホウレンソウは長日条件において,花芽形成と抽苔が誘導されます。
ホウレンソウでは、葉腋に集散花序 (cymose inflorescence)を持つ苞葉が求頂的(acropetal)に形成されます。
つまり、植物全体における花の配列順序としては,密錐花序 (thyrse inflorescence)ということになります。
生育状態にもよりますが、ホウレンソウの花は柄が短いために、複数の花が密集した花房として観察されます。
品種や系統によっては、雄株において茎頂に近い苞葉はあまり発達しない場合があるようです。
また、理由は分かっていませんが同じ条件で育成しても雌株よりも雄株の方が早く抽苔する傾向があります。
ホウレンソウの花



雄花は,4枚の萼片と4本の雄蕊(写真では葯しか見えていません)を持ちます。
雌花は,2枚の萼片と1本の雌蕊(写真では、1本の雌蕊の先端部が数本に分岐した形状の柱頭が写っています)を持つ。
写真で示した両性花は,雄ずいと雌ずいを1本ずつ持ちますが,これよりも多い本数の雄ずいを持つ両性花も見られます。
いずれの花も,花弁はありません。ホウレンソウは風媒花なので,訪花動物を誘引する役割を担う花弁は必要無いために退化したと考えられます。
植物の単性花(雄花または雌花)は、形態的特徴に基づいて、2つのタイプ(Type I および Type II ; Mitchell & Diggle 2005)に分けられています。
Type I : 雄花および雌花から,未発達の(または退化した痕跡的な器官として)雌蕊または雄蕊がそれぞれ観察される。
Type II : 雄花および雌花から,雌蕊または雄蕊の痕跡的器官が見出されない。(ホウレンソウの単性花は、このタイプ)
3つのクラスのホメオティック遺伝子によって両性花の形態形成機構を説明している"ABCモデル" に基づくならば,Type I の単性花は形成初期において雄蕊および雌蕊(心皮)の形成領域に相当するwhorl 3(クラス B とクラス C のホメオティック遺伝子が働く領域)およびwhorl 4(クラス C のホメオティック遺伝子のみが働く領域)の両方が確立されるが、一方の器官は途中で発達停止(退化)するために機能せず最終的に単性花になると解釈できま す。
Type IIに分類される単性花を形成する植物では、ホメオティック遺伝子の発現パターンの違いによってwhorl 3(雄蕊形成領域)およびwhorl 4(雌蕊[心皮]形成領域)のどちらか一方だけが確立される仕組みを持つ可能性が考えられます。実際、Type II の単性花を形成するホウレンソウにおいて、whorl 3(雄蕊の形成領域)の領域決定に関わるクラスBのホメオティック遺伝子の発現を雄花で確認できますが、雌花ではほとんど確認できないことが報告されてい ます。しかし、クラス B 遺伝子が性決定遺伝子であることを支持する証拠は今のところありません。おそらく、ホウレンソウのクラスB遺伝子は性決定遺伝子の制御下にあると推定され ます。
雄株・雌株・間性株について
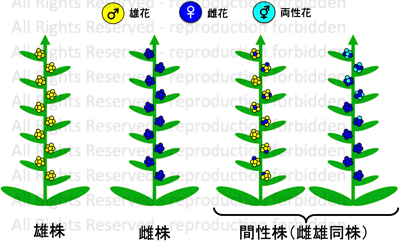
基本的に,雄株は雄花のみ、雌株は雌花のみを着生します。
その一方で,雌花と雄花または両性花を様々な比率で着生する間性株があります。
これまでに私たちは、間性の発現を支配する主働遺伝子が、雄決定遺伝子と連鎖していることを明らかにしてきました。
現在、間性主働遺伝子と雄決定遺伝子の単離(コード配列の同定)に取り組んでいます。
しかし、間性株には雌花率(1株あたりの雌花の着生割合)の連続的な変異が見出されるため、単一の主働遺伝子以外にも間性の発現に影響を及ぼす遺伝子があると推定してます。ホウレンソウの性決定機構を包括的に解明するには、まだ長い道のりが待っていそうです。
参考文献
Sherry RA, Eckard KJ and Lord EM, Flower Development in Dioecious Spinacia oleracea (Chenopodiaceae) (1993), American Journal of Botany 80: 283-291
西、平岡、ホウレンソウの性発現に関する研究(1961) 農業技術研究所報告 E 園芸 9: 129~159
Mitchell C, Diggle P. (2005) The evolution of unisexual flowers: morphological and functional convergence results from diverse developmental transitions. American Journal of Botany 92: 1068–1076.
Pfent C, Pobursky KJ, Sather DN, Golenberg EM, Characterization of SpAPETALA3 and SpPISTILLATA, B class floral identity genes in Spinacia oleracea, and their relationship to sexual dimorphism. (2005) Dev Genes Evol 215: 132–142 Sather DN, Jovanovic M. and Golenberg EM, Functional analysis
of B and C class floral organ genes in spinach demonstrates their role
in sexual dimorphism. (2010) BMC Plant Biology, 10:46
このページに掲載した文章・写真・図などの転載を禁じます。
このページに関するお問い合わせは下記まで
小野寺康之 onodera(at)abs.agr.hokudai.ac.jp