

アルミニウム集積植物に関する研究
酸性土壌は世界の氷圏以外の陸地面積の約30%を占めるといわれている。酸性土壌には植物に対する様々なストレスが存在し、生育できる植物種を制限する要因となっている。これらストレスのうち、多くの酸性土壌で大きな制限要因となっているのは酸性化によって土壌中での溶解度が高まるアルミニウム(Al)の過剰害である。多くの植物はAlを葉にほとんど集積することはない。これは多くの植物がAl毒性を回避するためにAl排除能を発達させており、植物体内にAlを輸送・貯蔵するためのシステムも無いためである。しかし、酸性度の強い土壌(すなわち土壌中のAl濃度が高い)に自生する植物には、葉に乾燥重ベースで数千~数万mg kg-1以上のAlを集積する種がしばしば存在する。これらの植物は「Al集積植物」といわれており、障害を受けることなくAlを体内に集積する機構を持っている。Al過剰ストレスがより厳しい条件(酸性硫酸塩土壌など)では、体内のAl集積能を高めることが生存においてより有利であるといわれている。我々はこのAl集積植物がもつ高Al耐性と高集積性に着目し、研究を行っている。
Al集積植物にはどのような種があるのだろうか?身近なところで茶樹やアジサイなどはAl集積植物としてよく知られている。茶樹の場合、私たちがお茶として飲んでいる茶葉は非常に若い葉であり、Al含有率は数百mg kg-1程度であるが、古い葉では30,000 mg kg-1を超えることがある。茶樹やアジサイなど、ほとんどのAl集積植物は木本種であり、多くは酸性土壌の多い熱帯を起源としているが、ソバのような冷涼な地域に生育する草本のAl集積植物もまれに存在する。ちなみに、これまでに報告されている中で最も葉のAl含有率が高い植物はSymplocos spicata(ハイノキの一種)という木本植物であり、葉のAl含有率は72,000 mg kg-1に達すると報告されている。この植物の体内でAlが水酸化物で存在すると仮定すると(実際の形態は未解明だが)、乾燥重の20%以上がAl化合物で占められることになる。18世紀のヨーロッパや、インドネシアでは現在でも、この植物の葉と樹皮をミョウバン(硫酸アルミニウムカリウム)の代わりとして染物に使っている(媒染剤)(図1)。そもそもミョウバンがなぜ染色に使われるかというと、含まれているAlイオンが植物染料に含まれる化合物とキレートを形成して鮮やかな色を呈すためである。同様の反応はAl集積植物であるアジサイの花(萼(がく)片)の中でも起こっている。アジサイの萼中でAlはdelphinidin 3-glucosideおよび3-caffeoylquinic acidと青色の複合体を形成し、萼の色を鮮やかな青にしているのである。アジサイを酸性度の低い土壌で生育させた場合、複合体形成に十分な量のAlを吸収することができず花の色が紫やピンクになってしまうことがある。アジサイ以外のAl集積植物でも青系の花を咲かせるものが多いといわれており、Al集積との関連性が議論されている。
図1. Symplocos spicata(右)とそれを使った染色(左)
植物界全体で葉に含まれる様々な元素の含有率を調べたとき、種間における含有率の変動が最も大きいのはAlである(図2)。同じ条件で育ててもAl集積植物は非集積植物の100倍以上のAlを葉に集積する。この差を決める要因は何なのであろうか?それを考える前にまずは植物界におけるAl集積植物の分布について考えてみよう。筆者はベルギーのルーベンカトリック大学(現ドイツ・ウルム大学)のJansen博士らとともに被子植物におけるAl集積植物の分布について詳しく調べている。数千種に及ぶ植物葉のAl含有率分析結果を解析したところ、Al集積植物は真性双子葉植物の進化的に新しい主要なクレード、すなわちバラ類やキク類に多く分布し、特にEricales(ツツジ目)、Gentianales(リンドウ目)、およびMyrtales(フトモモ目)において極めて多く存在することがわかった。一方、原始的被子植物ではAl集積植物はほとんど存在しなかった(図3)。このように植物の進化とAl吸収能には深い関係があると考えられ、後で述べる体内でのAlの形態の進化・変遷などもふまえて研究を進展させていきたい。
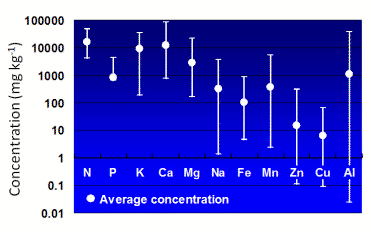
図2. インドネシアに生育する様々な植物種(308種、1174 サンプル)の葉に含まれる各元素の平均含有率及び変動範囲
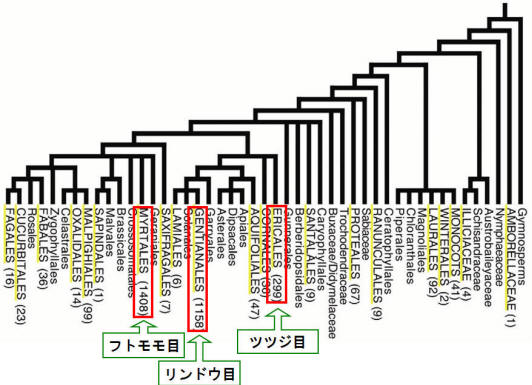
図3. 種子植物におけるAl集積植物の分布
大文字で記載したクレイドにはAl集積植物が見つかっており、括弧内の数は種数を示す。
次に、具体的にどのようなメカニズムでAl集積植物は体内のAlを無毒化しているのかを述べたいと思う。Al集積植物は以下の二つのAl毒性回避メカニズムを持つ。一つ目のメカニズムは細胞内でAlイオンを有機酸などのリガンドと結合させ、不活性化することである。基本的に、非Al集積植物が根からAlキレート化合物を分泌し、根圏においてAlを不活性化する機構と作用は同じであるが、根から分泌する場合と比べて体内では拡散が抑えられるため、より高濃度のAlイオンを不活性化することが可能である。また、土壌中に存在するAl結合生無機アニオン(ケイ酸やフッ素)を積極的に吸収し、それらとAlの複合体を体内で形成する植物も報告されており、これらの種では光合成産物をAl無毒化のための化合物に配分する必要が無いため、繁殖においてより有利であるかもしれない。もう一つのメカニズムは体内における隔離である。細胞レベルでは液胞やアポプラストにAlを隔離していることが報告され、細胞質における各種代謝が正常に保たれるようにしていると考えられる。組織レベルでは葉における表皮や、幹における樹皮のような植物体を外界から保護する役割を持つ部位で高濃度のAlが検出されている。これらの有害元素隔離はナトリウム集積植物や重金属集積植物でも類似して認められており、共通の過剰毒性回避メカニズムであるのかもしれない。
ここで我々が研究対象としているAl集積植物のひとつであるメラストーマ(Melastoma
malabathricum
L.、ノボタンの一種)を例に、Al集積植物とAlの関係について具体的に説明しよう。メラストーマは熱帯アジアの酸性硫酸塩土壌のような強酸性土壌の優占種の一つであり、葉に10,000
mg kg-1以上のAlを集積する。図4にこの植物におけるAlの輸送と集積の概略図を示した。まず根圏(根の近傍)に存在するAlイオンを根の細胞内に取り込む。Alイオンがどのように根の原形質膜を通過するのかはメラストーマも含めてAl集積植物では明らかにされていないが、0.5
mMのAlを含む培養液で栽培したメラストーマ根のAl吸収速度は新鮮重1
gあたり100~200
nmol h-1程度である。なお、後述のようにメラストーマ根はAlに対して親和性の高いゲル状多糖を分泌しており、このことがAl吸収を高める一因になっている可能性もある。Alが細胞内に取り込まれるのとほぼ同時に根のクエン酸合成酵素活性が急激に上昇し、根のクエン酸含有率はAl未処理の根と比べて数十~百倍以上になる。このクエン酸合成の誘導はAlに特異的である。この高濃度のクエン酸とAlイオンはキレート錯体を形成し、木部導管を通って葉に輸送されるが、養分状態が悪いなどの影響で根が十分な量のクエン酸を合成できない場合は、Alは地上部に運ばれない。すなわち、体内でのAl輸送能力の有無が地上部におけるAlの過剰集積に深く関係していることが予想される。葉に輸送されたAlは主として液胞中に集積すると考えられ、集積形態は輸送形態とは異なり単量体のAlイオンとシュウ酸とのキレート錯体である。地上部には輸送されず、根に集積するAlの形態もメラストーマではAl-シュウ酸キレート錯体である。

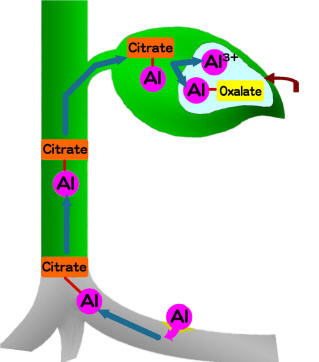
図4.
細胞内に入ったアルミニウムは合成が誘導されたクエン酸(citrate)とキレートし、導管を通って葉に輸送される。葉に到達したアルミニウムは液胞中にモノマーあるいはシュウ酸とのキレート形態して集積する。
メラストーマは非酸性土壌(土壌溶液中のAlイオン濃度が低い)でもAlを高集積できる。そのメカニズムについては根からのH+分泌による根圏pHの低下が主因と考えていたが、近年、別の興味深い新たなメカニズムを見つけた。メラストーマは土壌溶液中のAl濃度が低下すると大量のゲル状多糖類を根端から分泌し(図5)、このゲルはAlイオンのような3価カチオンに対して非常に親和性の高い吸着剤として働くのである(ちなみにトウモロコシの分泌するゲル状多糖は2価カチオンに対する親和性高い)。しかも、27Al NMRによる解析結果から吸着されたAlは無機モノマーの形態で非常にゆるくゲル中に保持・濃縮されており、濃縮されたAlイオンを根が吸収することを明らかにした。すなわち、メラストーマはAlを積極的に獲得するために、「Al欠乏」土壌ではゲル状多糖類を分泌し、根圏のAlイオンを濃縮、吸収しているのである(図6)。このような必須元素以外の無機元素吸収に関わる根分泌物はこれまで見つかっておらず、極めて興味深い。
 図5.
メラストーマの根が”Al欠乏”下で分泌するゲル状多糖類
図5.
メラストーマの根が”Al欠乏”下で分泌するゲル状多糖類
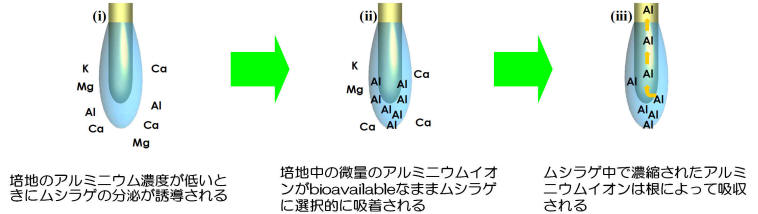
図6. メラストーマ根が分泌するゲルの機能の概略図
このように輸送時と集積時でキレーターを変えるメリットは何であろうか?メラストーマの導管液pHは約5であり、この条件下ではシュウ酸はカルシウムと難溶性沈殿を形成するため、カルシウム輸送が阻害されてしまう。このため、輸送時にはシュウ酸ではなくクエン酸をキレーターとして選んだと考えられる。Al輸送時にクエン酸をキレーターとするAl集積植物はメラストーマの他にもソバ、茶樹やアジサイが報告されている。ソバはメラストーマと系統樹ではかなり離れているが、集積形態は同じAl-シュウ酸複合体であり、Al集積メカニズムに関しては非常に類似している。しかしソバではAlによってクエン酸の合成が誘導されることが無く、恒常的に濃度が高いという点でメラストーマと大きく異なる
(5)。
最初に述べたように、多くの植物にとってAlイオンは有害である。では、なぜメラストーマのような植物は有害なAlを積極的に吸収するのか?実は生育に対するAlの有益性についても多くの報告があり、その研究の歴史はAl毒性の研究と同じぐらい古い。Alの有益性が報告されている植物種の多くは酸性土壌に適応した植物であるが、Al集積植物でその効果は特に顕著である。例えばアジサイ、茶樹、メラストーマ、Miconia albicans(ノボタン科)などはAlにより生育が促進されると報告されている。我々もAl集積植物の生育におけるAlの有益性については注目している。標準培養液を用いた水耕実験でメラストーマの根を観察すると、Alを培養液に添加していない場合は根は黒みを帯び、細根も少ないが、Alを添加すると根は白色になり、細根も増加する。また、根のリグニン含量はAl無添加で栽培した場合に高く、何らかのストレスをAl不在下で受けており、Alによりそれが改善されることは間違いない。Al不在下で生育させたメラストーマでは、葉においても変色や丸まりなどの異常な症状が確認され、Al添加により正常になる(図7)。これらのAlの有益な効果を説明するものとして、AlによるH+毒性の軽減やリン酸過剰の軽減が提案されているが、メラストーマも含めた多くの植物種の生育促進はこれらの仮説では説明できない。

図7. 標準培養液で長期間栽培したメラストーマの葉
培養液にアルミニウムを添加しない場合、葉の丸まりなど形態的に異常な症状が見られる(左)。培養液に0.5
mMの塩化アルミニウムを添加した場合は正常な形態である(右)。
我々のこれまでの研究で、メラストーマの生育においてなぜAlが有益に働くかを説明する理由として、二つのことを明らかにしている。一つめはAl不在時におけるCa等必須元素の体内での不活性化である。前述の通り、メラストーマは体内でのAl集積におけるリガンドとしてはシュウ酸を、輸送におけるリガンドとしてはクエン酸を使っている。クエン酸についてはAl添加によって合成が大きく誘導されるが、シュウ酸に関しては根、地上部ともに恒常的に濃度が高い(図8)。ご存知のとおり、シュウ酸は多くの多価カチオンと不溶性の塩を作ってしまう(例えばCa。シュウ酸Caは人間の結石の原因でもある)。このため、Alが体内に存在すればシュウ酸はAlとキレートすることにより「無毒化」されるが、Alがない場合は他のCaのような多価カチオンと結合し、それらの欠乏を引き起こしてしまうのである。土壌溶液中にAlイオンが多量に存在する条件が一般的な生育条件であるメラストーマにとってAlイオンが無い条件はむしろ異常であり、Al無毒化機構が生育に対して負に働いたのである。
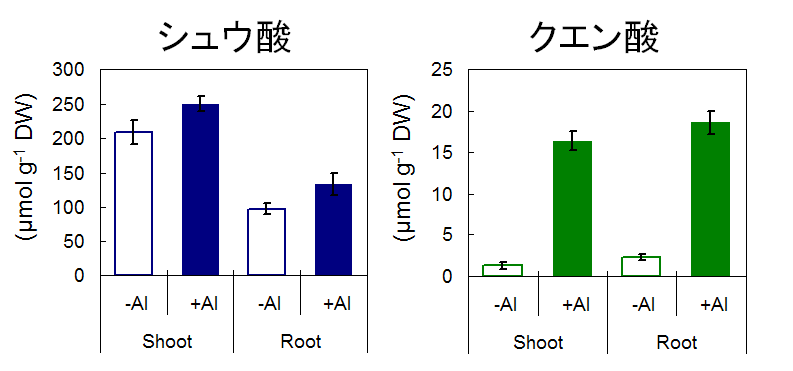
図8. ±0.5 mM Al処理した時のメラストーマの有機酸含有率
もう一つの理由は、メラストーマの環境適応に関係している。メラストーマは極めて土壌酸性度の強い酸性硫酸塩土壌の優占種の一つである。酸性硫酸塩土壌はパイライト(FeS2)などが酸化されることにより硫酸を生じ、土壌を強酸性化している。通常、好気的条件では土壌中の鉄イオン濃度は低いが、酸性硫酸塩土壌ではFeイオンの濃度もある程度まで高まる。実はメラストーマはこのFeイオンに対する耐性が極めて弱いのである。メラストーマはFeイオンにより酸化ストレスを受け、根は黒化し、生育が抑制される。しかし、そこにAlイオンを添加するとFeイオンによるストレスは解消される(図9、10)。これはメラストーマがAlイオンを過剰吸収することによってFeイオンの吸収を抑制しているためであることが明らかになった。すなわち、メラストーマが酸性硫酸塩土壌のような強酸性土壌で優占種となるためには、Alの超集積は必要不可欠な性質なのである。(つづく)
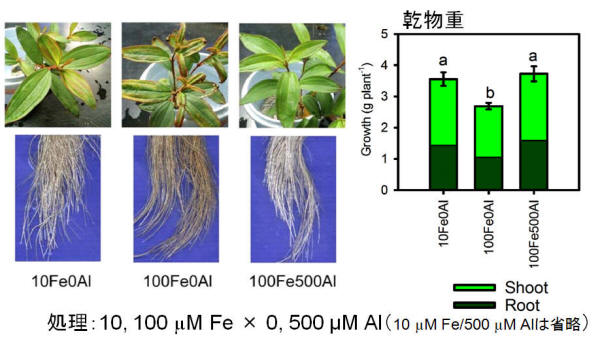
図9. Al×Fe処理を施したメラストーマの写真と乾物重
Feによる障害がAlによって解消されている。
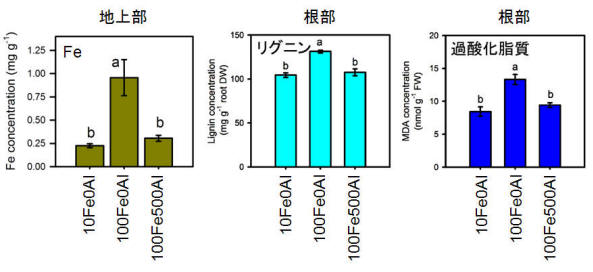
図10. Al×Fe処理を施したメラストーマのFe含有率、リグニン含有率、過酸化脂質レベル
AlによりFe吸収は抑制され、根のリグニンおよび過酸化脂質レベルは正常値に戻った。